我的购物车
我的购物车我的祖源成分
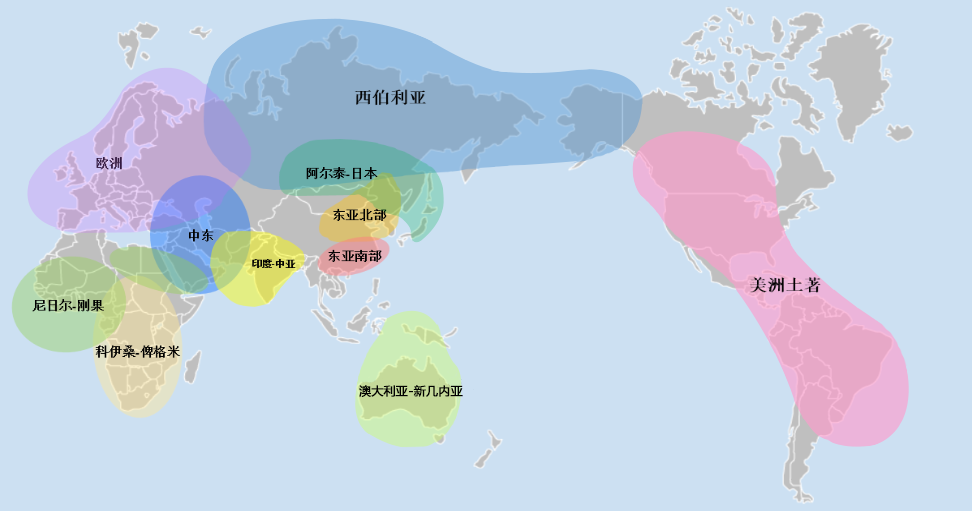
*地图上的色块,展示了该成分分布较多的地理范围
点击成分名称了解更多
东亚北部 55%
该成分是现代汉族的主体之一——北方汉族的特征成分,最多分布于中国华北地区的北方汉族,新疆维吾尔自治区锡伯族,内蒙古自治区达斡尔族和蒙古族,东北三省及北京、内蒙古、新疆一带的满族。此外,该成分在日本人群及朝鲜人群均有较高分布。
典型比例:北方汉族56%(汉藏语系-汉语族),蒙古族55%(阿尔泰语系-蒙古语族),日本49%,朝鲜族56%,纳西族52%(汉藏语系-藏缅语族),畲族46%(苗瑶语族系),彝族54%(汉藏语系-藏缅语族-彝缅语支),土族52%(阿尔泰语系-蒙古语族),土家族44%(汉藏语系-藏缅语族)。
收起东亚南部 20%
该成分是现代汉族的主体之一——南方汉族的特征成分,最多分布于中国华南地区的南方汉族,湘西地区的土家族、苗族,在亚洲东南部的阿美族、傣族比例可高达70%。
典型比例:南方汉族45%(汉藏语系),苗族40%(苗瑶语系),土家族33%(汉藏语系-藏缅语族),彝族19%(汉藏语系-藏缅语族-彝缅语支),纳西族19%(汉藏语系-藏缅语族),畲族32%(苗瑶语系),土族12%(阿尔泰语系-蒙古语族),日本12%,朝鲜族10%,蒙古族9%(阿尔泰语系-蒙古语族)。
收起阿尔泰-日本 13%
该成分最多分布于阿尔泰山脉东北部的西伯利亚地区及大兴安岭黑龙江流域,在日本岛也有较多分布。在中国东北地区的达斡尔族、赫哲族、鄂伦春族的比例约占18%,在日本人中比例约占26%。
典型比例:日本26%,乌尔奇人26%(阿尔泰语系-通古斯语族),赫哲族19%(阿尔泰语系-通古斯语族),鄂伦春族19%(阿尔泰语系-通古斯语族),锡伯族13%(阿尔泰语系-通古斯语族),朝鲜族14%,汉族8%(汉藏语系-汉语族),蒙古族13%(阿尔泰语系-蒙古语族)。
收起美洲土著 8%
该成分是美洲印第安人的特征成分,在现存的印第安人里比例可高达90%。该成分最多分布于墨西哥南部的玛雅人,墨西哥南方哈瓦那州西部的米斯特克人以及亚利桑那希拉河和索尔特河沿岸的皮马人。
典型比例:玻利维亚88%,哥伦比亚24%,玛雅人86%(玛雅语系),墨西哥44%,朝鲜族1%,日本2%,蒙古族4%(阿尔泰语系-蒙古语族),汉族2%(汉藏语系-汉语族)。
收起西伯利亚 3%
该成分是西伯利亚地区的特征成分,最多分布于北极地区的爱斯基摩人,西伯利亚东北部的楚科奇人,俄罗斯勘察加半岛南部的勘察加人,俄罗斯勒那河以东的尤卡吉尔人,以及散居在克拉斯诺亚尔斯克北部埃文基和泰梅尔民族区的雅库特人。
典型比例:爱斯基摩人67%(爱斯基摩-阿留申语系),阿留申人19%(爱斯基摩-阿留申语系),勘察加人74%(楚科奇-勘察加语系),尤卡吉尔人58%(尤卡吉尔语系),汉族占5%(汉藏语系-汉语族),蒙古族4%(阿尔泰语系-蒙古语族),朝鮮族9%,日本5%等。
收起印度-中亚 1%
该成分最多分布于印度各主要民族,在古吉拉特人、泰卢固人、旁遮普人、孟加拉人中比例约占60%,在巴基斯坦北部兴都库山脉东部的卡拉什人及南亚的泰米尔人中比例也较高。
典型比例:古吉拉特人60%(印欧语系-印度伊朗语族)、泰卢固人64 %(达罗毗荼语系-东南语族)、旁遮普人55 %(印欧语系-印度伊朗语族)、孟加拉人56%(印欧语系-印度伊朗语族)、卡拉什人42%(印欧语系-印度伊朗语族),泰米尔人63%(达罗毗荼语系-南部语族),朝鲜族1%,日本人0%,蒙古族1%(阿尔泰语系-蒙古语族),汉族0%(汉藏语系-汉语族)。
收起中东 1%
该成分最多分布于伊朗中部的波斯人,土耳其及西欧等国家和地区的土耳其人,中东地区的游牧民族库尔德人。聚居在埃及尼罗河流域的埃及人也有较多分布,比例高达63%。
典型比例:伊朗50%(印欧语系-印度-伊朗语族),埃及63%(亚非语系-闪米特语族),贝多因人62%(亚非语系-闪米特语族),塞浦路斯人57%(印欧语系-希腊语族)、德鲁兹人63%(亚非语系-闪米特语族),沙特阿拉伯人72%(亚非语系-闪米特语族),朝鲜族0%,日本人0%,蒙古族4%(阿尔泰语系-蒙古语族),汉族0%(汉藏语系-汉语族)。
收起欧洲 0%
该成分是欧洲的特征成分,最多分布于欧洲北部的挪威人、芬兰人、白俄罗斯人,欧洲南部的希腊人和西班牙人,欧洲西部的法国人,欧洲东部的乌克兰人、匈牙利人及德系犹太人。
典型比例:法国57%(印欧语系-罗曼语族),英国65%(印欧语系-日耳曼语族),西班牙48%(印欧语系-罗曼语族),意大利37%(印欧语系-罗曼语族),匈牙利60%(乌拉尔语系芬兰-乌戈尔语族),德系犹太人28%,乌克兰人62%(印欧语系-斯拉夫语族-东斯拉夫语支),捷克人61%(印欧语系-斯拉夫语族-西斯拉夫语支),朝鲜族0%,日本人0%,蒙古族0%(阿尔泰语系-蒙古语族),汉族0%(汉藏语系-汉语族)。
收起澳大利亚-新几内亚 0%
该成分是大洋洲土著居民的特征成分,最多分布于太平洋西部新几内亚岛及其附近岛屿上的土著民族,比例可高达99%。
典型比例:巴布亚人99%(巴布亚诸语言),朝鲜族1%、日本人3%,蒙古族2%(阿尔泰语系-蒙古语族),汉族1%(汉藏语系-汉语族)。
收起科伊桑-俾格米 0%
该成分是非洲中部和南部的特征成分,主要是分布在非洲中西部的热带雨林狩猎采集族群。最多分布于非洲南部喀拉哈里盆地的Juǀ'hoan及ǂKhomani族群,在非洲中西部热带雨林以狩猎采集为生的姆布蒂人和比亚卡人也有较多分布,分布在东非坦桑尼亚的哈扎人该成分的比例也较高。
典型比例:北Juǀ'hoan 85.50%(科伊桑语系),姆布蒂人76.20%(尼罗-撒哈拉语系或尼日尔-刚果语系),ǂKhomani 72.16%(科伊桑语系),比亚卡人59%(尼日尔-刚果语系),哈扎人 49.48%,朝鲜族1%,日本0%,蒙古族1%(阿尔泰语系-蒙古语族),汉族0%(汉藏语系-汉语族)。
收起尼日尔-刚果 0%
该成分是非洲西部的特征成分,在东非沿海也有较多分布,与非洲西部地区的农民扩张有关。最多分布于非洲西海岸的门德人、南非班图人及肯尼亚班图人,分布于西非尼日利亚西部和西南部的约鲁巴人也有较高比例,同时在赤道以南、东非沿海的卢希亚族群里比例也较高。
典型比例:约鲁巴人72%(尼日尔-刚果语系),门德人69%(尼日尔-刚果语系),卢希亚人61%(尼日尔-刚果语系),埃桑人72%(尼日尔-刚果语系),基库尤人53%(尼日尔-刚果语系),肯尼亚班图人61%(尼日尔-刚果语系),朝鲜族1%,日本人0%,蒙古族2%(阿尔泰语系-蒙古语族),汉族0%(汉藏语系-汉语族)。
收起祖源成分如何得出?
源基因提供的祖源成分解读,是根据源基因用户的基因数据和地理、民族信息,以及1000 Genomes Project等基因组数据库,使用美国加州大学洛杉矶分校(UCLA)研发的Admixture祖源分析软件推导得出的,共涉及11类祖源成分。将您在源基因测序获得的常染色体DNA数据和不同地区种族人群标准参考数据库进行比较,从而模拟出由哪些人群按何种比例混合而成。
关于祖源成分,你还要了解这些
所有关于个人基因来源比例的分析都不是完全客观的,而是严重依赖于参考样本的选取。举例说,如果参考样本里面只有“蒙古族”、“日本人”、“泰国人”而没有“汉族”,并将聚类分析得到的成分以这三个群体来命名,那么一个汉族人得到的成分结果就很可能是这三个成分的混合,而不会得到一个称作“汉族人”的来源。这并不意味着汉族是由这三个群体混合得来的,而只是这些族群在一些基因成分上有共同来源,但并不说明传播方向。不同版本的祖源成分计算器选用的位点、参考序列以及处理流程不同,同样的测试结果在不同的计算器推导的结果也可能不同。 某种祖源成分的名称只是指该成分在现代的该地区(或族群)中高频出现,如您和该地区共享某一成分,并不明确表示您的基因来自该地区,而仅仅表示您和该地区的群体的相似程度。
50元无门槛优惠券会以邮件或短信的形式自动发送填写调查问卷即可获得50元源基因产品优惠券
参与研究
中国是一个多民族聚居和融合的国家,历史上人口曾经多次迁移和融合,近现代也有大量人口流动。由于国内目前还没有较好的祖源参考数据库,祖源成分分析结果还不够细致和精准。我们希望借助自己的基因组数据库,通过一份简单的有偿祖源信息调查,构建一个庞大、准确且丰富的中国人祖源分析系统。 为了帮助构建祖源分析系统,我们需要您提供本人、父母及祖父母的姓氏、民族、出生地等信息。随着数据的积累和方法的更新,源基因祖源分析的准确性和多样性将会不断提升。我们期待你可以告诉我们更加全面准确的祖源信息,你的参与至关重要。
点击填写祖源信息参考文献
- Skoglund P, Thompson J C, Prendergast M E, et al. Reconstructing Prehistoric African Population Structure[J]. Cell, 2017, 171(1):59-71.
- Lutz M W, Crenshaw D, Welsh-Bohmer K A, et al. New Genetic Approaches to AD: Lessons from APOE-TOMM40, Phylogenetics[J]. Current Neurology & Neuroscience Reports, 2016, 16(5):1-9.
- Sudmant P H, Rausch T, Gardner E J, et al. An integrated map of structural variation in 2,504 human genomes[J]. Nature, 2015, 526(7571):75-81.
- Rasmussen M, Guo X, Wang Y, et al. An Aboriginal Australian Genome Reveals Separate Human Dispersals into Asia[J]. Science, 2011, 334(6052):94-8.
- Mills R E, Klaudia W, Chip S, et al. Mapping copy number variation by population scale genome sequencing[J]. Nature, 2011, 470(7332):59-65.
- Kato M, Kawaguchi T, Ishikawa S, et al. Population-genetic nature of copy number variations in the human genome[J]. Human Molecular Genetics, 2010, 19(5):761-773.
- Jakobsson M, Scholz S W, Scheet P, et al. Genotype, haplotype and copy-number variation in worldwide human populations[J]. Nature, 2008, 451(7181):998-1003.
- Kim J J, Verdu P, Pakstis A J, et al. Use of autosomal loci for clustering individuals and populations of East Asian origin[J]. Human Genetics, 2005, 117(6):511.
 3.12
3.12